БУДЕТ ЛИ СЛЕДУЮЩАЯ "ЗЕЛЕНАЯ РЕВОЛЮЦИЯ"?
Доктор биологических наук В. СОКОЛОВ (г. Новосибирск)
ТРИУМФ КАРЛИКОВ
В 1970 году Норману Борлаугу присудили Нобелевскую премию мира за так называемую "зеленую революцию". В чем же она заключалась? К середине XX века сельское хозяйство получало огромное количество минеральных удобрений, но существующие сорта растений не могли эффективно трансформировать их в урожай зерна. Из-за высоких концентраций питательных веществ в почве злаки быстро росли, набирали зеленую массу, а затем полегали, что существенно снижало намолоты. При этом индекс урожая (отношение веса зерна к общему весу наземной массы) был значительно ниже 50 процентов, то есть основным продуктом оказывались солома и листья (даже в пересчете на сухое вещество).
Для борьбы с полеганием Борлауг предложил использовать растения с коротким стеблем. Признак короткостебельности достаточно просто контролируется генетически и легко передается через гибридизацию. Полученные Борлаугом полукарликовые сорта также формируют большую наземную массу, но уже за счет высокой кустистости, при этом не полегают и дают хороший урожай с индексом около 50 процентов. Кроме того, эти сорта обеспечивают более эффективное использование удобрений.
Растения обычных сортов вначале накапливают соединения азота в зеленой массе, а затем после цветения переносят их в зерновки. Короткостебельные сорта отличаются тем, что восстанавливают и переносят азот до тех пор, пока не закончится налив семян. Усвоение азота из почвы у них продолжается много дольше и приводит к большей продуктивности. Благодаря "зеленой революции" Мексика за 15 лет увеличила производство пшеницы в три раза (на аналогичный прирост Европе потребовалось 150 лет) и из крупнейшего импортера превратилась в экспортера зерна.
Залог успеха "зеленой революции" в том, что базовый признак, на основе которого строилась селекционная технология, имеет простой генетический контроль и со 100-процентной вероятностью передается потомкам. Именно это позволило создать и поддерживать необходимую на практике устойчивую генетическую систему.
НЕЗАМЕЧЕННАЯ СЕНСАЦИЯВ конце прошлого века наиболее сенсационным научным событием стало, несомненно, клонирование овцы в Эдинбурге (Великобритания). Нет ничего удивительного в том, что эти эксперименты взбудоражили общество: результат, полученный на млекопитающих, впечатляет уже потому, что его можно явно или неявно примерить к человеку. Овечка Долли настолько приковала к себе всеобщее внимание, что на этом фоне в России остались незамеченными сообщения об "апомиктической революции" и ее перспективах, появившиеся в западных популярных и научных изданиях. Приведем заголовки лишь некоторых из них: "Апомиксис: социальная революция сельского хозяйства" ("Biotechnology & Development Monitor", 1994); "Апомиксис: бесполая революция" ("Science", 1996); "Революция в производстве гибридной кукурузы" ("Agricultural Research", 1998); "Никакого секса, пожалуйста, мы - растения" ("Economist", 1997); "Огромное будущее апомиксиса" ("Trends in Plant Science", 1998).
Из приведенных заголовков видно, что как научные, так и популярные издания, обсуждая прикладное значение генетически регулируемого апомиксиса, используют термин "революция". Иными словами, все признают глубину преобразования агротехнологий в случае практического освоения данного способа репродукции растений.
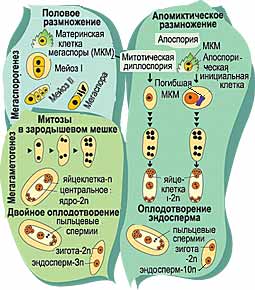
Что же такое апомиксис? Это специфический способ размножения, позволяющий получать абсолютные генетические копии материнского растения, то есть созданная природой технология клонирования. Чтобы понять, в чем заключается принципиальное значение апомиксиса для селекции, начнем со сравнительного анализа полового и апомиктичного типов размножения.
ЗАЧЕМ РАСТЕНИЯМ ПОЛОВОЕ РАЗМНОЖЕНИЕ?Наиболее часто встречающийся и знакомый всем половой способ размножения сельскохозяйственных растений связан с циклическим чередованием спорофитной и гаметофитной фаз. Спорофит - обычная, наблюдаемая нами форма растений, несущая двойной набор хромосом. Гаметофитная фаза у цветковых растений весьма коротка и характеризуется одинарным (гаплоидным) набором хромосом. Напомним, что хромосомы - это структурные элементы ядер клеток, в которых содержится ДНК, несущая практически всю генетическую информацию. Каждый вид растений имеет строго определенное число и характерную форму хромосом. В световой микроскоп хромосомы можно увидеть только тогда, когда клетка начинает готовиться к делению и ДНК конденсируется с помощью белков.
Специальные клетки в органах размножения, имеющие двойной набор хромосом, проходят клеточное деление особого типа - мейоз. Образуются микроспоры (мужские клетки) и мегаспоры (женские клетки), которые несут одинарный хромосомный набор. Эти клетки снова делятся, но уже по механизму митоза, при котором число хромосом в дочерних клетках не изменяется. Из микроспоры формируется трехъядерная пыльца, а из мегаспоры - восьмиядерный зародышевый мешок, заключенный в материнскую семяпочку. После оплодотворения образуются диплоидный зародыш и триплоидный эндосперм.
Замечательное следствие этих событий в том, что набор хромосом во вновь образованных спорофитах снова диплоидный, но генотип каждого потомка (совокупность генов) качественно отличается от родительских и между собой, то есть каждый потомок уникален и непохож на родителей.
Зачем нужны такие довольно длинные пути создания и объединения половых продуктов? Казалось бы, проще размножаться почкованием, чем тратить энергию и строительные материалы на формирование органов полового размножения, тем более, что всегда есть риск, что растение окажется неопыленным, то есть не оставит потомства и выбудет из борьбы за существование. Однако смысл в том, что живым организмам необходимо постоянно адаптироваться к изменяющимся условиям среды, то есть иметь запас вариабельности, что и достигается через комбинативную изменчивость, сопровождающую половое размножение.
Так, например, у ячменя имеется 7 пар хромосом, поэтому при формировании гаплоидных половых клеток возможны 128 (27) вариантов их комбинаций. В свою очередь при оплодотворении возможное число вариантов потомков будет равно числу сочетаний из 128 по 2, что обеспечивает огромный резерв изменчивости.
Кроме того, размножение через семенную фазу в отличие от вегетативного служит мощным барьером против клеточных инфекций. Вспомните, сколько проблем доставляет картофель, который поражают более 500 видов внутриклеточных паразитов.
ЧТО ДИКОМУ ХОРОШО, ТО КУЛЬТУРНОМУ - СМЕРТЬЕсли в дикой природе комбинативная изменчивость -необходимый компонент устойчивости и приспособляемости вида, то у культурных сортов она нежелательна, так как разрушает ценные комбинации генов, контролирующих хозяйственно-полезные признаки, по крупицам собранные поколениями селекционеров. Поэтому существует настоятельная необходимость уйти от половой репродукции, и одна из возможностей сделать это - бесполосеменное размножение (апомиксис). При апомиксисе жизненный цикл укорочен, а эмбриогенез осуществляется в результате деления неоплодотворенных клеток, не прошедших через редукцию (стадию уменьшения) числа хромосом. При этом происходит передача полного материнского генотипа следующему поколению, то есть клонирование.
В основном у растений встречаются два типа апомиксиса - диплоспория и апоспория. При диплоспории зародышевый мешок развивается из материнской клетки мегаспоры, не прошедшей мейоза, и таким образом неоплодотворенная яйцеклетка, которая далее дает начало следующему поколению, воспроизводит генетическую копию материнского растения. При апоспории зародыш формируется из соматической клетки и также является абсолютным генетическим клоном.
Отсюда можно заключить, что бесполосеменное размножение позволяет поддерживать генетически стабильный клон через семенное размножение неограниченно долго. Если добиться наследствен но контролируемого превращения половых растений в апомиктические, то мы революционизируем селекционные технологии. Так, например, гетерозисные (высокопродуктивные) гибриды экономически очень выгодны. Однако их использование рентабельно только тогда, когда от одного акта опыления получают не менее 500-1000 семян. По этим причинам гетерозис успешно реализован на практике лишь у многосемянных культур (кукуруза, томаты, огурцы). Основные хлебные злаки и рис, несмотря на многочисленные попытки, остались за чертой гибридной селекции; между тем решение этой проблемы позволило бы увеличить производство на 15-30 процентов на уже освоенных площадях.
Многие дикие растения апомиктичны от природы (одуванчик, мятлик, лапчатка), однако среди культурных видов их единицы (некоторые кормовые травы, малина, земляника). У читателя может возникнуть вопрос: если апомикты имеют столько замечательных качеств, почему они оказались вне практического использования? Ответ прост: именно отсутствие необходимой для совершенствования растений вариабельности не позволяло отбирать среди них ценные для селекции варианты. Еще в самом начале окультуривания растений человек неосознанно работал с материалом, способным выщеплять необходимые для улучшения признаки, что позволяло пересортировывать их, используя половое размножение, и таким образом совершенствовать агрономические качества. Апомиксис как раз исключает эту возможность. В свое время отсутствие расщепления в опытах на апомиктичной ястребинке заставило Менделя усомниться во всеобщем характере принципов гибридологического поведения признаков, открытых им на горохе. По этой же причине только в самое последнее время удалось селекционно улучшить некоторые апомиктичные кормовые травы.
ПЕРВОПРОХОДЕЦ АПОМИКСИСА - КУКУРУЗАПервый в мире проект по созданию апомиктичного коммерческого сорта у культурных растений был объявлен и начат в СССР в 1958 году. Его автор - профессор Д. Ф. Петров, создавший для выполнения данного исследования лабораторию цитологии и апомиксиса растений в только что организованном Институте цитологии и генетики Сибирского отделения Академии наук. Однако идея закрепления гетерозиса через размножение, исключающее сегрегацию генов, принадлежит замечательным ученым, сотрудникам Н. И. Вавилова, - М. С. Навашину и Г. Д. Карпеченко, с которыми Петров работал перед войной в знаменитом ВИРе (Всесоюзном институте растениеводства). Получение апомиктов планировалось на важнейшей сельскохозяйственной культуре - кукурузе.
В работе, начатой Д. Ф. Петровым, использовали несколько подходов, некоторые из которых сейчас кажутся наивными, но они вполне соответствовали научным представлениям того времени. Один из них оказался продуктивным и продолжается в России в нашей лаборатории, а также в США, во Франции, в Мексике до настоящего времени. В этом исследовании передачу кукурузе (Zea mays L. - Zm) апомиктического способа репродукции осуществляли от ее дикого сородича - трипсакума (Tripsacum dactyloides L. - Td) путем гибридизации. Предполагалось, что бесполосеменное разможение у трипсакума контролируется двумя генами: один препятствует редукции числа хромосом, другой стимулирует развитие эмбриона из яйцеклетки, минуя опыление. Отсюда можно заключить, что для передачи такого типа размножения кукурузе необходимо перенести максимум две хромосомы от трипсакума в случае расположения предполагаемых генов в разных хромосомах. Считалось, что присутствие столь незначительного генетического материала от дикого родителя не окажет решающего влияния на внешний вид и хозяйственно-полезные признаки апомиктичной кукурузы.
На основе таких допущений и начались скрещивания кукурузы с ее диким родственником - трипсакумом. К удаче исследователей, оказалось, что признак бесполосеменного размножения передается по наследству получаемым гибридам.
Д. Ф. Петров и его коллеги создали целый ряд апомиктических гибридных линий. Наиболее стабильными из них были 38-хромосомные (20Zm+18Td), то есть содержащие 20 хромосом кукурузы и 18 хромосом трипсакума. Но они из-за большого количества генетического материала от дикого родителя оказались далеки от кукурузы, что исключало перспективу их коммерческого использования.
НА ПОРОГЕ НОВОЙ "ЗЕЛЕНОЙ РЕВОЛЮЦИИ"Исходя из знаний генетики кукурузы, мы могли надеяться на достаточно быстрое "совершенствование" гибридов в сторону культурной формы. Американские генетики к середине 90-х годов прошлого столетия показали, что культурная кукуруза отличается от дикой единичными генами, контролирующими развитие (по разным оценкам, от 3 до 5). Поэтому, проводя отбор среди естественных и искусственно получаемых мутантов, можно достаточно просто усилить агрономически ценные признаки у бесполосеменных гибридов. Используя гамма-лучи в качестве мутагенного фактора, сотрудник нашей лаборатории Б. Ф. Юдин получил "кукурузоподобные" 58-хромосомные гибриды, весьма близкие к культурному родителю. Эти растения, как и кукуруза, имели по одному стеблю и по 12-14 рядов в початке. К сожалению, после обработки гамма-лучами признак апомиксиса проявлялся нестабильно, но стало очевидным, что совершенствование агрономичес ких признаков у гибридов реально.
В 1993 году мы получили предложение Министерства сельского хозяйства США провести совместные исследования по апомиктичной кукурузе с использованием нашего экспериментального материала. Кроме того, проект поддержал Российский фонд фундаментальных исследований. Работа была начата весной 1994 года в Оклахоме (США). Среди привезенных из Новосибирска линий гибридов сразу же удалось выявить несколько 39-хромосомных апомиктов. Примечательно то, что все они имеют идентичный набор из девяти одних и тех же хромосом трипсакума. Именно эти девять хромосом минимально необходимы для поддержания апомиктического способа размножения у гибридов. К сожалению, "дикие" хромосомы довольно существенно ухудшали хозяйственно-важные признаки культуры, прежде всего вес семян. У гибридов он был в среднем равен 0,06 г, что гораздо ближе к весу семян трипсакума (~0,03 г), чем кукурузы (~0,22 г). Кроме того, с точки зрения коммерческой перспективы была нежелательна избыточная кустистость гибридов.
Понимая, что апомиктичные гибриды должны будут конкурировать за рынок с кукурузой, мы провели сравнительный анализ признаков, по которым они ее превосходят. К таковым необходимо отнести урожай зеленой массы, высокое содержание в ней протеина и других переваримых компонентов, содержание в семенах полиненасыщенных жирных кислот, устойчивость растений к засухе, переувлажнению и засолению почвы.
Эти несомненные преимущества позволили бы уже сейчас использовать гибриды в качестве фуражной культуры. Однако пока они не могут давать семян из-за полной мужской стерильности (апомиктам необходимо оплодотворение центральной клетки, иначе зерновки не развиваются). Чтобы получить потомство от гибридов, приходится рядом высевать кукурузу в качестве опылителя. У первоначально выделенных 39- и 49-хромосомных линий фертильность (количество способных к дальнейшему развитию семян) была не более 3-5 процентов на початок. Нам удалось выяснить, что в значительной степени это связано с особенностями роста пыльцевых трубок при опылении. Подбором опылителей и других факторов удалось добиться 50-процентной фертильности, что с учетом большого числа початков на растениях-гибридах позволяет им конкурировать по семенной продуктивности с кукурузой.
Наша апомиктичная кукуруза уже запатентована в США и в 11 других странах. Но самое главное, создана огромная коллекция апомиктичных линий, которую можно будет использовать в качестве исходного материала как в академических исследованиях, так и в практической работе по созданию коммерческих сортов.
Сейчас стоит вопрос, кто первым в мире создаст апомиктичное культурное растение и реализует коммерческий сорт с использованием этого типа размножения. Двадцать лет назад нам казалось, что это будет сделано в нашей стране. Десять лет назад у нас появились сразу два сильных конкурента - Вэйн Ханна, работающий с жемчужным просо в США, и Ив Савидан, очень быстро повторивший наш путь гибридизации кукурузы с трипсакумом (совместный проект Международного института пшеницы и кукурузы в Мексике). Активно работают по генетике апомиксиса на модельных объектах Австралия и Европейский союз, постоянно привлекая российских ученых, к сожалению, лишь на правах исполнителей.
Сможем ли мы при столь значительной конкуренции сохранить позиции в дальнейшем? Науку делают не только идеи, но и методы, требующие значительных материальных ресурсов, а наши ресурсы несопоставимо меньше, чем у конкурентов. Достаточно долго нам удавалось, имея существенный задел, сохранять лидерство. Однако его легко потерять, и в данном случае навсегда, так как все, сделанное после первого апомиктического сорта, будет только повтором. Итак, вопрос о новой "зеленой революции" пока открыт, но до ее начала остается немного времени. Кто же будет первым? Вопрос в науке не праздный, ибо, как считал П. Л. Капица, быть лидером в науке или идти, пусть на полшага, но позади - принципиально разные состояния. Сможет ли российская наука адекватно отреагировать на вызов времени или мы будем с грустью вспоминать былые успехи и то, что Россия была духовной матерью второй "зеленой революции"? Здесь возможно повторение ситуации, случившейся с первой "зеленой революцией". Работа по созданию неполегающих пшениц на хорошо удобренных почвах была начата и успешно развивалась в 30-е годы XX века в Италии. Однако итальянским исследователям начатую работу не удалось довести до мирового признания, прежде всего из-за ее невостребованности и изоляции страны при Муссолини, а после войны идея была реализована Н. Борлаугом.
Доктор биологических наук В. СОКОЛОВ (г. Новосибирск).Автор благодарен Российскому фонду фундаментальных исследований (грант 00-04-49542) за поддержку данной работы.
Читайте в любое время
