Крошечные стражи подземных галактик
Марина Павлова, Всероссийский научно-исследовательский институт биологической защиты растений. Фото автора
Слой почвы, непосредственно прилегающий к поверхности корней и называемый ризосферой, обильно заселён микроорганизмами.
Статья — победитель конкурса научно-популярных статей, проходившего в рамках XIII Всероссийской олимпиады «Нанотехнологии — прорыв в будущее!», организованной МГУ им. М. В. Ломоносова и ФИОП (группа РОСНАНО).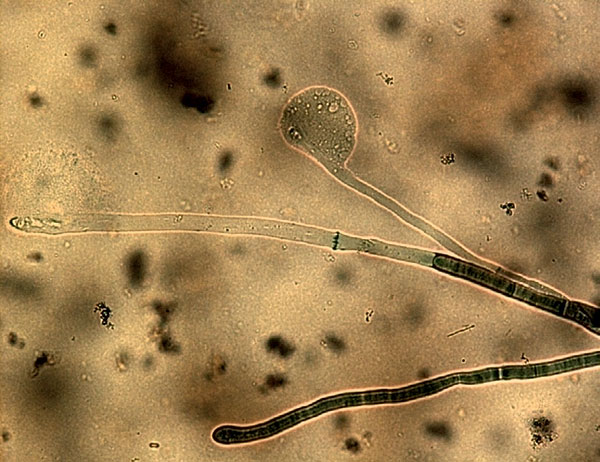
Корневые системы растений занимают в почве огромные пространства. Суммарная длина корней пшеницы без учёта корневых волосков, по разным данным, может достигать от 60—70 до 600 км! Если же учитывать корневые волоски (выросты клеток покровной ткани корня в зоне поглощения), которых на одном растении может быть до 14—15 млрд, то суммарная длина корней может достигнуть 10 000 км! Суммарная поверхность корней ржи, подсчитанная в одном из исследований, опять же без корневых волосков, составила 237 м2. Это примерно площадь теннисного корта.
Слой почвы толщиной около 2—5 мм, непосредственно прилегающий к поверхности корней и называемый ризосферой (от греч. rhiza — корень и sphaira — шар, сфера), обильно заселён микроорганизмами. Размеры бактерий составляют в среднем 0,5—3 мкм (10-6 м), поэтому для них корневая система с указанной площадью поверхности составляет если не галактику, то целый континент размером, скажем, с Австралию. Естественно, что поверхность других органов растения — стеблей, листьев (филлосфера), цветов, плодов, семян (спермосфера) — также населяют микроорганизмы, но такого их обилия и разнообразия, как в прикорневой зоне, нет.
Ризосфера значительно отличается по своим свойствам от «свободной» почвы. Здесь на 1—2% выше влажность, несколько другая реакция среды (обычно если почва кислая или щелочная, то ризосфера ближе к нейтральной), выше содержание органических веществ, растворимость некоторых минеральных веществ, например соединений железа и марганца. Ризосфера — лакомая экологическая ниша (вернее, совокупность множества мини-экологических ниш) для почвенных обитателей — от микроскопических бактерий и грибов (и даже водорослей!) до червей всех типов и членистоногих.
Что же их так привлекает в этом узком участке почвы? В первую очередь корневые выделения. Физиология растений такова, что до 20—25% усвоенного ими в процессе фотосинтеза углерода теряется с корневыми выделениями. Причём это не только пассивные потери, а по большей части активная секреция корнем различных веществ. То есть корень не только орган поглощения воды и минералов, но и орган синтеза и переработки веществ. Например, органические кислоты, выделяемые корнем, повышают растворимость соединений фосфора, кальция и железа в почве, делая их более доступными для усвоения растением. Многие низкомолекулярные соединения корневых выделений могут подавлять рост других растений-конкурентов или фитопатогенов. Эти вещества называют фитонцидами.
Есть вещества, которые выделяются при естественном отмирании корней, «слущивании» корневых чехликов и т. д.
Несмотря на кажущуюся расточительность растений, такие траты вполне оправдывают себя. Для микроорганизмов ризосфера благодаря корневым выделениям — источник легкоусвояемых органических веществ. Это, прежде всего, сахара — глюкоза, фруктоза, мальтоза и др., и органические кислоты — например яблочная, лимонная, янтарная, а также аминокислоты, витамины и др. Всё это очень быстро поглощается и перерабатывается для своих нужд теми представителями почвенной микробиоты, которые оказались самыми «расторопными» и сумели заселить прикорневые зоны быстрее других.
Но какой толк от этого растению? Если в почве небольшое содержание спор и клеток болезнетворных грибов и бактерий (невысокий инфекционный фон), то при благоприятных погодных условиях ризосфера растения заселяется преимущественно дружественными микроорганизмами. Теми, которые не агрессивны к нему, не претендуют на его клетки и ткани, а, наоборот, в обмен на «стол и дом» оказываются полезными ему в меру своих не столь малых, как может показаться, сил. Такие микроорганизмы, которые стимулируют рост растений, называются PGPR-бактериями (Plant Growth Promoting Rhizo bacteria). Наиболее часто встречаются PGPR-штаммы среди родов Psеudomonas, Bacillus, Azospirillum, Azotobacter, Clostridium, Enterobacter, Klebsiella и др.
Агенты PGPR, как и любые другие супер-агенты, обладают суперспособностями. Они быстро и эффективно заселяют (колонизируют) ризосферу, интенсивно размножаются и увеличивают свою численность в ней; устойчивы к неблагоприятным факторам внешней среды — колебаниям температуры, рН почвы, влажности и т. д. Эти микроорганизмы усваивают не только легко доступные корневые выделения, но и разнообразные питательные субстраты; при этом они улучшают минеральное питание растения-хозяина, защищают его от патогенных микроорганизмов.
PGPR не только быстро заселяют ризосферу, но и успешно удерживают завоёванные позиции от посягательств других колонизаторов. Например, фитопатогенных грибов, которые при контакте с корнем разрушают его покровы, проникают во внутренние ткани (паренхиму), поражают проводящую систему (сосуды и ситовидные трубки) и способны распространяться выше по стеблю вплоть до генеративных органов (цветов и плодов). Фитопатогенные грибы повинны, например, в таких заболеваниях корней растений, как корневые гнили, «чёрная ножка», «снежная плесень», полегание сеянцев, увядание и т. д. Корни, поражённые фитопатогенными грибами, темнеют, становятся склизкими, легко разрушаются, приобретают неприятный запах. Всё это объясняется действием ферментов и токсинов грибов, разрушающих клеточные стенки и мембраны растительных клеток. Высвобождающееся содержимое клеток становится субстратом для развития и жизнедеятельности патогенов. Поэтому их называют зловещим словом — некротрофы, то есть питающиеся мёртвым.
Интересно, что при выращивании на искусственных питательных средах фитопатогенные грибы выглядят очень красиво. Например, возбудители корневых гнилей пшеницы — грибы рода Fusarium на искусственной питательной среде (картофельно-глюкозный агар) в чашке Петри выглядят как пушистый бело-розовый мех. Если же в чашку Петри совместно с грибом высеять культуру каких-либо бактерий, то зачастую мицелий гриба даже «не заметит» её и полностью заполнит чашку. А вот если в одной чашке Петри выращивать мицелий гриба вместе с культурой PGPR, то рост фитопатогена в какой-то момент останавливается, и между культурой бактерий и мицелием гриба образуется «зона отчуждения» — полоса питательной среды, не занятая ни одним из противников. Её называют антифунгальной зоной.
Дело в том, что PGPR-агенты выделяют вещества, разрушающие гифы фитопатогенных грибов, причём изначально стратегия PGPR направлена на «изгнание» конкурентов. Если рассмотреть под микроскопом гифы грибов при совместном культивировании с PGPR, то можно наблюдать явное повреждение мембран и растворение внутреннего содержимого гиф. Иногда можно увидеть разрыв кончика гифы с выходом содержимого наружу.
Всё это — результат действия антифунгальных (противогрибковых) антибиотиков, выделяемых PGPR. В большинстве своём они обладают свойствами биосурфактантов, то есть веществ, снижающих поверхностное натяжение на границе раздела фаз, в том числе на поверхности клеточных мембран, в связи с чем нарушается стабильность грибной клеточной мембраны — плазмалеммы. Известно также, что многие антибиотики PGPR негативно воздействуют на эргостерол — основной компонент грибных мембран, что обусловливает избирательность действия, так как у животных, например, основной компонент клеточных мембран — холестерол.
Большинство PGPR — не облигатные (необязательные) симбионты высших растений, поэтому помимо корневых выделений и отмерших частиц корней они способны поглощать и перерабатывать большое количество различных субстратов. В том числе и своих конкурентов за ризосферу. То есть PGPR могут просто съесть мицелий фитопатогенных грибов. Так, на седьмые—десятые сутки совместного культивирования фитопатогена и PGPR можно увидеть скопление бактериальных клеток на поверхности гиф с последующим их полным разрушением.
Установлено, что помимо антибиотиков большинство PGPR-бактерий синтезируют также ферменты, расщепляющие компоненты грибной клеточной стенки — хитин в комплексе с β-глюканами и белковый матрикс (из гликопротеинов). Расщепление остальных компонентов грибных клеток не составляет труда для широкого круга ферментов PGPR.
Такое «поедание» гиф вызывает удивление и восхищение, однако трудно сказать, происходит ли это в реальных природных условиях. Как жаль, что невозможно (ну или пока невозможно) установить на корнях микроскопическую видеокамеру и понаблюдать за взаимоотношениями ризосферных жителей, подобно тому, как мы можем наблюдать жизнь животных во влажных тропических лесах.
Вообще, микробные сообщества ризосферы очень динамичны, состав их меняется по мере роста и развития растения. Так, например, в период активного роста растения в ризосфере могут доминировать представители рода Pseudomonas, хорошо усваивающие органические кислоты в выделениях молодых корней, а по мере старения растения начинают преобладать бактерии рода Bacillus, более эффективно утилизирующие вещества отмирающих корешков.
Однако, даже если PGPR-агенты функционировали в ризосфере лишь непродолжительное время, они так или иначе оставляют свой след в жизни растения. Особенно интересно, что помимо прямого подавления конкурентов в ризосфере PGPR могут достаточно тонко воздействовать на растения опосредованно. Например, путём повышения его иммунитета. Дело в том, что большинство PGPR, как мы могли убедиться, довольно агрессивны в смысле завоевания жизненного пространства и использования пищевых субстратов. Заболевания у растения они не вызовут, но могут быть им распознаны как потенциальные патогены. То есть вызовут у растения реакцию, подобную реакции человека на прививку. У растений этот отклик называется индуцированной системной устойчивостью. Проявляется она в усиленной лигнификации (одревеснении) тканей корня и в синтезе фитоалексинов — соединений, выделяемых растением при заражении и запускающих каскад реакций, направленных на уничтожение патогенов. Таким образом, растение, заселённое PGPR, становится вооружённым и готовым к возможному нападению.
Помимо этого практически все PGPR, оправдывая своё имя, в той или иной мере стимулируют рост растений за счёт выделения растительных гормонов: ауксинов, цитокининов, гиббереллинов и подобных им веществ — в небольших, но значимых количествах. Благодаря этому растение быстрее «проскакивает» ювенильный (юношеский) период, когда оно наиболее уязвимо для патогенов. Надо отметить, что сами патогены также часто синтезируют фитогормоны, причём в огромных дозах, с целью выращивания растительной биомассы для собственного потребления. В результате у растения формируются уродливые наросты, галлы, курчавости, «ведьмины мётлы» и т. д. Кстати, растительные гормоны гиббереллины были впервые выделены не из растений, а как раз из фитопатогенных грибов Gibberella fujikuroi, вызывающих заболевание «бешеный рис» — чрезмерное удлинение стеблей риса и практически полное отсутствие зерна.
Ещё одно важное свойство PGPR, как и любых других прокариот, — способность к выделению широкого спектра ферментов и биологически активных веществ — гораздо более широкого, чем у эукариот. Поэтому они могут переводить минеральные элементы, необходимые для питания растений (например, фосфор или железо), в доступную для них форму. Также многие PGPR обладают способностью к азотфиксации — усваиванию атмосферного азота и включению его в органические вещества — аминокислоты и белки. Частью этих соединений PGPR делятся с растением-хозяином. Этот процесс называется ассоциативной азотфиксацией. Интересно, что в настоящее время способность к азотфиксации обнаружена у большинства прокариот, а не только у широко известных клубеньковых бактерий Rhizobium.
Итак, PGPR обладают многими потенциально полезными качествами для растений. Однако их всё же нельзя рассматривать как абсолютных супергероев, всю свою жизнь посвящающих борьбе с фитопатогенами и самоотверженной защите растений. Просто для них наиболее выгодная экологическая стратегия — это формирование дружественной ассоциации с растением и поддержание его сильным, здоровым и продуцирующим корневые выделения как можно дольше. По своей значимости для растения сообщества PGPR в ризосфере можно сравнить со значимостью микрофлоры здорового кишечника для человека. Точно так же, как и PGPR, обитатели кишечника (в основном бифидо- и лактобактерии) оберегают своё жилище от посягательств других микроорганизмов, в том числе болезнетворных, участвуют в переваривании сложных органических веществ, особенно целлюлозы (клетчатки), синтезируют витамины, незаменимые аминокислоты и др.
В последние годы, подобно тому, как медицина уделяет большое внимание нормализации кишечной микрофлоры человека, в сельском хозяйстве ведётся множество исследований, направленных на поддержание «здоровой микрофлоры» в почве. Такую почву называют супрессивной, то есть подавляющей развитие растительных патогенов естественным образом.
PGPR-агенты активно используются в биологической защите растений в виде микробиопестицидов. В России наиболее известны микробиопестициды «Фитоспорин» на основе Bacillus subtilis, штамм 26Д, «Псевдобактерин» на основе Pseudomonas aureofaciens, «Ризоплан» на основе Pseudomonas fluorescens, штамм АР-33 и др. По существу, эти препараты представляют собой жидкие культуры PGPR-бактерий (бактериальные клетки в растворе питательной среды). Ими обрабатывают, например, семена перед посевом (вместо препаратов под страшным наименованием «протравители»). С поверхности семян PGPR-агенты распространяются на корни прорастающего семени, где успешно функционируют с обоюдной пользой для себя и для растения. Однако «работают» такие препараты только при соблюдении севооборотов, низком инфекционном фоне в почве, благоприятных погодных условиях и правильном применении. Пока что в большинстве хозяйств это труднодостижимые идеалы, и потому микробиопестициды используются относительно редко и считаются не очень эффективными. Однако положительные сдвиги в применении технологий биологической защиты растений есть, интерес к ним растёт с каждым годом, а значит, PGPR и технологии их применения, возможно, станут основой «природного земледелия» будущего.
Словарик
Экологическая ниша — место обитания вида в природе, включающее как положение его в пространстве, так и функциональную роль в сообществе, отношение к абиотическим условиям существования. Понятие экологической ниши описывает не столько место пребывания организмов, сколько их «профессию». Например, в ризосфере это может быть утилизация слущивающихся клеток корневого чехлика или потребление субстратов на стыке ветвления корней.
Микробиота — совокупность микроорганизмов того или иного местообитания. Синоним микрофлоры, однако многие считают этот термин устаревшим и неправомерным из-за того, что бактерии и другие микроорганизмы уже давно не относят к флоре. Однако в данной статье оба термина равнозначны.
Штамм — чистая культура вирусов, бактерий, других микроорганизмов или культура клеток, изолированная в определённое время и в определённом месте. Той или иной совокупностью свойств PGPR обладают именно определённые штаммы, например Bacillus subtilis 26Д, а не весь вид Bacillus subtilis целиком.
Гифы (от др.-греч. ὑφή — паутина) — нитевидные образования, из которых состоит мицелий грибов. Могут быть многоклеточными или одноклеточными, но со множеством ядер. Основная функция гиф — поглощение воды и питательных веществ.
Прокариоты — живые организмы, не обладающие оформленным клеточным ядром и другими внутренними мембранными органоидами (хлоропластами, митохондриями, эндоплазматической сетью и др.). Включают два домена (надцарства): бактерии и археи.
Эукариоты — домен (надцарство) живых организмов, клетки которых содержат ядро.
Читайте в любое время
